Ганглии вегетативной нервной системы локализация свойства. Ганглии нервной системы
Подробности
Ганглии представляют собой скопления мультиполярных (один аксон и несколько дендритов) нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия.
Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны . В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля 1-го типа), чувствительные (клетки Догеля П-го типа) и ассоциативные (клетки Догеля Ш-го типа) . Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки 1-го типа называют длинноаксонными нейронами. Нейроны П-го типа - равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки П-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля Ш-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
Ацетилхолин
- никотиновые (блок кураре, гексаметоний), мускариновые (блок атропин) рецепторы. Активация рецепторов→генерирование ВПСП. Быстрый ВПСП (N-холиноцер)→открытие ионных каналов. Медленный ВПСП (М-холинорец)→подавление М-тока, обусловленного повышением К-проводимости.
Нейропептиды
– действуют как нейромодуляторы.

Энкефалины
, вещество Р, люлиберин, нейротензин, соматостатин – симп. ганглии (+Ach)
Катехоламины
(NA, дофамин) – нейромедиаторы мелких клеток с интенсивной флуоресценцией.
Нейропептид Y, соматостатин – симп. постганглионары.
Симпатические постганглионары: NA, АТФ, нейропептид У.
α1→инозотолтрифосфат, диацилглицерол. α2→активация G-белка, ↓цАМФ.
β→G-белок→AC→цАМФ
Исключения
: медиатор Ach, рецепторы мускариновые.
Парасимп. постганглионары: Ach, ВИП, NO, соматостатин, АТФ, опиоидные пептиды.
М1 (высокое сродство к пирензепину)-повышение секреции кислоты клетками желез желудка, М2 (низкое)- замедляют серд. ритм, секреция слезных и слюнные желез.
Разнообразное действие:
-Специфические втор. посредники: М2 может акт-ть IP3, а может инг-ть AC, уменьшая цАМФ.
-Действие на К и Са-каналы
-На эндотелии образуется NO→гуанилатциклаза→цГМФ→ цГМФ-зависимая протеинкиназа→расслабление глажких мышц.
Вегетативная нервная система (ВНС) в основном обеспечивает иннервацию внутренних органов.
Делится на:
Симпатический отдел
Парасимпатический отдел
Метасимпатический (Энтеральный)
Отличия вегетативной нервной системы от соматической НС:
- Не находится под контролем сознания
- Возможность автономного функционирования (даже при полном нарушении связи с ЦНС)
- Генерализованный характер распространения возбуждения в периферическом отделе ВНС (особенно в симпатическом отделе).
- Наличие вегетативного ганглия в эфферентном отделе рефлекторной дуге. Таким образом, эфферентная часть ВНС представлена двумя нейронами: преганглионарный нейрон в пределах ЦНС (ствол мозга, спинной мозг), постганглионарный нейрон в вегетативном ганглии. Т.е. тела последних нейронов вегетативных дуг вынесены за пределы ЦНС.
- Низкая скорость проведения нервного импульса (преганглионарные волокна типа В, постганглионарные типа С)
- Ткани-мишени для ВНС: гладкомышечные клетки, поперенополосатая сердечная мышца, железистая ткань (для соматической — поперечнополосатая скелетная МТ). Симпатические волокна способны влиять на гликогенолиз в печени и липолиз в жировых клетках (метаболический эффект)
Обычно внутренние органы имеют двойную иннервацию: симпатическую и парасимпатическую, однако мочевой пузырь и ресничная мышца получают в основном парасимпатическую, сосуды, потовые железы, волоковые мышцы кожи, селезёнка, матка, мозг, органы чувств, надпочечники – только симпатическую.
Высшие вегетативные центры
структуры лимбической системы, базальные ганглии, КГМ, гипоталамус (передние ядра – зона парасимпатических ядер, задние — зона симпатических ядер), центральное серое вещество среднего мозга, ретикулярная формация (её нейроны формируют жизненно важные центры продолговатого мозга ССЦ, ДЦ).
Нервные центры (центральный отдел) симпатической нервной системы – промежуточно-латеральные ядра боковых рогов спинного мозга C VIII — L II — III
Нервные центры (центральный отдел) парасимпатической нервной системы – вегетативные ядра III пары (глазодвигательный нерв – Ядро Якубовича), VII (лицевой нерв – верхнее слюноотделительное), IX (языкоглоточный нерв – нижнее слюноотделительное), X (блуждающий нерв – заднее ядро), промежуточно-латеральные ядра спинного мозга S II -S IV
На уровне рабочих отделов заложены эфферентные клетки, аксоны которых идут не сразу к рабочему органу, в отличие от соматических, а прерываются в периферическом вегетативном ганглии. Здесь они переключается на последние нейроны, Волокна нейронов спинного мозга называются преганглионарными. Преганглионарные волокна переключаются в вегетативном ганглии на следующий нейрон, аксон которого носит название постганглионарного.
Симпатический вегетативный ганглий
Ганглий покрыт сверху капсулой. Здесь имеются следующие клетки:
- Чувствительные нейроны
- Эфферентные нейроны
- Хромаффинные клетки, выделяющие катехоламины (регулируют уровень возбудимости клеток узла.
Функции ганглия: проводниковая, замыкательная и рецепторная.
Нейроны вегетативного ганглия обладают теми же свойствами, что и нейроны центральной нервной системы.
Парасимпатический вегетативный ганглий
Ганглий покрыт сверху капсулой. В нём имеются следующие клетки:
- Чувствительные — клетки Догеля 2-го типа, их рецепторы могут быть механо-, термо-, хемочувствительными.
- Эффекторные нейроны – клетки Догеля 1-го типа, имеют много коротких дендритов и один аксон, уходящий за пределы ганглия.
- Вставочные – клетки Догеля 3-го типа.
- В ганглии имеются также хромаффинные клетки, выделяющие катехоламины, возможно, серотонин, АТФ, нейропептиды (регуляторная функция).
Физиология вегетативного ганглия
(переключение с преганглионарных волокон на постганглионарные)
- Низкая лабильность нейронов вегетативных ганглиев (10-15 импульсов в секунду), у соматических 200 имп/сек.
- Длительная синаптическая задержка, больше в 5 раз.
- Большая длительность ВПСП (20-50 мс), длительность потенциала действия 1,5-3 мс из-за продолжительной следовой гиперполяризации нейронов ганглия.
- Большую роль имеет пространственная и последовательная суммация.
- Медиатор: в вегетативных ганглиях – преганглионарные нейроны выделяют АХ.
- На уровне ганглия хорошо развита конвергенция и дивергенция (мультипликация).
Симпатический отдел вегетативной нервной системы
Симпатические вегетативные ганглии расположены в симпатическом стволе, превертебральных узлах, узлах сплетений (брюшное аортальное, верхнее и нижнее подчревное).
Преганглионарные волокна короткие, сильно ветвятся. Постганглионарные волокна длинные тонкие многократно ветвятся, образуют сплетения. Хорошо развита мультипликация.
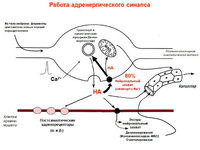
Медиатор постганглионарных адренергических симпатических волокон – НА (90%), адреналин (7%), дофамин (3%). Медиатор стойкий, длительно проявляет свою активность. НА связывается с α и β-адренорецепторами органов-эффекторов. Классификация основана на чувствительности их к фарм.препаратам: α-адренорецепторы блокируются фентоламином, β — пропранололом. Адренорецепторы имеются не только на органах, иннервируемых симпатическими волокнами (сердце, жировая ткань, сосуды, мышца-дилататор зрачка, матка, семявыносящий проток, кишечник) (α 1 и β 1), но и вне синапсов (на тромбоцитах, скелетные мышцы, эндокринные и экзокринные железы) (α 2 и β 2), а также на пресинаптческой мембране.
Передача возбуждения осуществляется быстрее, чем по симпатическому отделу. Влияния краткосрочные.
Влияния:
- Постоянное (тоническое)
- Фазное (пусковое) – резкое изменение функции (зрачковый рефлекс)
- Адаптационно-трофическое
Адапционно-трофическое влияние симпатической нервной системы Орбели-Гинецинского
Это приспособление обменных процессов к уровню функциональной активности. Идею о трофическом влиянии сформулировал И.П.Павлов. В опыте на собаке обнаружил симпатическую веточку, идущую к сердцу, раздражение которой вызывало усиление сердечных сокращений, без изменения частоты. Усиление сокращений утомлённой мышцы связано с активацией обменных (трофических) процессов под влиянием НА. Он активирует специфические рецепторы мембраны мышечных волокон, запускает каскад химических реакции в цитоплазме, ускоряющих синтез макроэргов, повышает возбудимость периферических рецепторов. Предполагается наличие трофогенов в нервных окончаниях. К трофогенам относятся нуклеотиды, некоторые аминокислоты, простагландины, катехоламины, серотонин, АХ, сложные липиды, ганглиозиды.
Парасимпатическая отдел вегетативной нервной системы
Парасимпатические вегетативные ганглии (далеко от ЦНС) расположены внутри органов (интрамурально) или околоорганно (ресничный, крылонёбный, ушной, подъязычный, поднижнечелюстной узлы), в узлах сплетений.
Преганглионарные волокна длинные, слабо ветвятся. Постганглионарные волокна короткие, мало ветвятся. Мультипликация развита слабо.
Медиатор постганглионарных парасимпатических волокон АХ.
Ацетилхолин на клетках-эффекторах связывается М-холинорецепторами. М-холинорецепторы возбуждаются мускарином, блокируются ядом кураре.
Ацетилхолин – нестойкий медиатор, основная часть разрушается ацетилхолинэстеразой до холина и ацетата, которые затем захватываются пресинаптической мембраной и используются для синтеза. Меньшая часть диффундирует в интерстиций и кровь.
Влияния:
- Постоянное (тоническое)
- Фазное (пусковое) – резкое изменение функции (торможение работы сердца, активация перистальтики, сужение зрачка)
Тонус вегетативных центров
Многие преганглионарные и ганглионарные нейроны обладают постоянной активностью, называемой тонусом. В покое частота электрических импульсов в вегетативных волокнах 0,1-5 имп/с. Тонус вегетативных нейронов подвержен суточным колебаниям: симпатотонус днём выше, ночью ниже, во время сна повышается тонус парасимпатических волокон. Симпатотонус обеспечивает постоянный тонус сосудов. Тоническое влияние блуждающего нерва (ваготонус) на сердце постоянно сдерживает ЧСС. Чем выше двигательная активность у человека, тем больше выражен парасимпатический тонус (уменьшение ЧСС у спортсменов). Причины вегетативного тонуса:
- Спонтанная активность. Высокий уровень спонтанной активности характерен для нейронов РФ.
- Поток афферентных импульсов от различных рефлексогенных зон.
- Действие БАВ и метаболитов
Вегетативные рефлексы. Классификация:
По уровню замыкания:
- центральные (соматовегетативный рефлекс- имеет общую афферентную часть с соматическим рефлексом)
- периферические, автономные (дуга рефлекса может замыкаться вне ЦНС в вегетативном ганглии интраорганно или экстраорганно, возможно существование аксон-рефлекса)
По расположению рецепторов:
Интероцептивные (механо-, хемо-, термо-, ноце-, полимодальные рецепторы)
а) Висцеро-висцеральные (каротидный синус, солнечное сплетение, перистальтика)
б) Висцеро-кутанные (соответственно зонам Захарьина-Геда)
в) Висцеро-моторные (раздражение интерорецепторов может вызывать моторные реакции).
- 4. Парасимпатический отдел внс, его центры, ганглии, медиаторы, внутриклеточные посредники, характер влияния на органы и ткани; регуляция активности синапсов.
- 1. Рефлекторный принцип деятельности цнс. Схема дуги соматического спинального рефлекса.
- 2. Открытие и.М.Сеченовым торможения в цнс. Виды и механизмы центрального торможения.
- 3. Роль спинного мозга в регуляции тонуса мышц и движений.
- 4. Симпатический отдел внс. Его центры, ганглии, медиаторы, внутриклеточные посредники, влияния на деятельность внутренних органов, регуляция активности синапсов.
- 1. Взаимоотношения между рефлексами в цнс. Принцип общего конечного пути.
- 2. Пресинаптическое торможение в цнс, его механизмы, значение.
- 3. Роль продолговатого и среднего мозга в регуляции тонуса мышц. Тонические рефлексы мозгового ствола.
- 4. Надсегментарные центры регуляции вегетативных функций. Гипоталамус как высший подкорковый центр регуляции вегетативной нервной системы.
- 1. Понятие о нервном центре. Основные свойства нервных центров.
- 2. Постсинаптическое торможение в цнс, его виды, механизмы, значение.
- 3. Роль мозжечка в регуляции тонуса мышц и движений.
- 4. Общий план строения вегетативной нервной системы, её отличия от соматической.
- 1. Виды центральных нейронов, их основные функции.
- 2. Явление суммации в нервных центрах. Виды и механизмы суммации.
- 3. Понятие о контрактильном тонусе. Децеребрационная ригидность, рефлекторный механизм её развития.
- 4. Синапсы вегетативной нервной системы, их виды, локализация, механизм возбуждения, основные механизмы регуляции деятельности синапсов.
- 1. Понятие о сегментарных и надсегментарных отделах цнс. Спинальный шок, причины и механизмы его развития.
- 2. Реципрокная иннервация мышц-антагонистов, её механизмы, значение.
- 3. Понятие о тонусе мышц. Виды тонуса. Основные принципы его поддержания. Этапы становления тонуса в онтогенезе.
- 4. Синапсы вегетативной нервной системы, их виды, локализация, механизм возбуждения, основные механизмы регуляции деятельности синапсов.
- 1. Эфферентная функция центрального нейрона. Место формирования распространяющегося возбуждения, виды импульсной активности нейронов.
- 2. Принцип доминанты в деятельности цнс. Свойства доминантного очага. Значение доминанты для интегративной деятельности организма.
- 3. Понятие о пирамидной и экстрапирамидной системах регуляции тонуса мышц и движений.
- 4. Вегетативные ганглии, их свойства. Понятие о метасимпатической нервной системе и ее медиаторах.
- 1. Рефлекс как основной принцип деятельности цнс. Основные этапы учения о рефлексе. Обратная афферентация, её значение для организма.
- 2. Первичное и вторичное торможение в цнс. Понятие о тормозных нейронах и синапсах.
- 3. Роль базальных ганглиев мозга в регуляции тонуса мышц и движений.
- 4. Схема дуги спинального вегетативного рефлекса; медиаторы
- 1. Интегративная деятельность центрального нейрона, её механизмы.
- 2. Основные принципы и механизмы координационной деятельности цнс.
- 3. Проприорецепторы, их роль в регуляции тонуса мышц, регуляция активности проприорецепторов.
- 4. Периферические вегетативные рефлексы, их дуги, значение для регуляции вегетативных функций.
4. Вегетативные ганглии, их свойства. Понятие о метасимпатической нервной системе и ее медиаторах.
Особенностью периферического звена вегетативной нервной системы является наличие ганглиев, представляющих собой скопление нейронов.
Вегетативные ганглии играют важную роль в распределении и распространении нервных влияний на органы. Отмечено, что число нервных клеток в ганглиях в несколько раз превышает число преганглионарных волокон.
В ганглиях наблюдается явление конвергенции. Вместе с этим обнаруживается явление пространственной и временной суммации. У вегетативных ганглиев проявляются те же свойства, что и у соматических нервных центров. Поэтому ганглии вегетативной нервной системы иногда называют нервными центрами, вынесенными на периферию.
Метасимпатическая (интраорганная) нервная система (МНС) представляет собой комплекс нервных образований – нейронов, тела которых формируют ганглии, и выходящих за пределы ганглия отростков нервных клеток. Эти структуры локализуются в стенке сердца, кишечника и других органов. Число нейронов этой системы превышает таковое в спинном мозгу. МНС не имеет центрального отдела, т. е. относительно автономна; ее функциональный модуль включает водитель ритма, сенсорные клетки, вставочные, тонические и эффекторные нейроны. Эти нервные образования обеспечивают автономию органов и местную регуляцию функций гладких и сердечной мышц, секреторного эпителия, аппарата всасывания и мелких кровеносных сосудов. Роль метасимпатической нервной системы особенно велика в регуляции функций кишечника (выше прямой кишки), где центральные нервные влияния практически отсутст вуют. В синапсах МНС обнаружено около 20 медиаторов и модуляторов, среди них ацетилхолин, холецистокинин, энкефалины, гистамин, серотонин, соматостатин, АТФ, вещество Р, катехоламины. Симпатические и парасимпатические нервы могут образовывать синапсы на метасимпатических нейронах и влиять на их активность.
Билет №8
1. Рефлекс как основной принцип деятельности цнс. Основные этапы учения о рефлексе. Обратная афферентация, её значение для организма.
Рефлекс (Р.) – это закономерная реакция организма на изменения внешней или внутренней среды, протекающая при участии нервной системы в ответ на раздражение рецепторов. Рефлекторная дуга – нервный путь рефлекса – состоит из чувствительного нервного окончания (или рецепторной клетки), чувствительного нервного волокна с ганглием, центральной части (чувствительных, вставочных, эффекторных нейронов разных уровней ЦНС), эфферентного нервного волокна и эффектора.Основоположником учения о рефлексе как реализуемой нервными центрами спинного мозга ответной реакции на раздражение явился французский философ, математик и физиолог Рене Декарт (1648). Он сформулировал два важных положения рефлекторной теории: 1) деятельность организма при внешнем воздействии является отраженной (впоследствии ее стали называть рефлекторной: лат. reflexus – отраженный); 2) ответная реакция на раздражение осуществляется при помощи нервной системы.Термин «рефлекс» впервые применил чешский физиолог, анатом и офтальмолог И. Прохазка, а выражение «рефлекторная дуга» – английский невропатолог и физиолог М. Холл. Новым шагом в развитии учения о рефлексе стала книга И. М. Сеченова «Рефлексы головного мозга» (1863),главной идеей которой явилось утверждение: «Все акты сознательной и бессознательной жизни суть рефлексы». Иными словами, И. М. Сеченов использовал рефлекторный принцип для объяснения механизмов деятельности головного мозга, в том числе процессов мышления. Отсутствие в ряде случаев видимой ответной реакции на действие стимулов ученый объяснил развитием центрального торможения, открытого им годом раньше(1862). Таким образом, рефлексы могут иметь «усеченный конец».И. П. Павлов, не будучи прямым учеником И. М. Сеченова, считал,однако, его своим учителем и высоко оценивал значение книги И. М. Сеченова, назвав ее «гениальным взмахом русской мысли».
И. П. Павлов более 30 лет своей жизни посвятил изучению высших рефлексов головного мозга, используя для этой цели метод условных рефлексов и, следовательно, объективный подход к изучению функций мозга. Он развил рефлекторную теорию, основанную на трех принципах: 1) детерминизма, т. е. причинной обусловленности различных процессов мозговой деятельности; 2) анализа и синтеза раздражений в высших отделах мозга; 3) приурочения динамики к структуре, т. е. связи функций мозга с определенными его структурами.Современный этап развития учения о рефлексе может быть назван системно-кибернетическим и связан с именами советских физиологовН. А. Бернштейна и П. К. Анохина. Идеи обратной связи были использованы для объяснения механизмов рефлекторной деятельности Н. А. Бернштейном(1947), в результате появился термин «рефлекторное кольцо».
П. К. Анохин(1949) назвал обратные связи рефлексов «обратной афферентацией» . Источником ее являются рецепторы, локализованные в органе-эффекторе (1) и в органах чувств, принимающих участие в оценке результата рефлекторного акта (2).Так, при исполнении какой-либо мелодии на музыкальном инструменте такими рецепторами могут быть проприорецепторы мышц и сухожилий руки (1), а также рецепторы органа слуха (2). Сигналы обратной афферентации используются для сравнения результата рефлекторного акта с его программой.
В естественных условиях жизнедеятельности рефлексы обычно объ-
единены в системы. Причем системообразующим фактором является общий результат, к которому приводит осуществление этой совокупностирефлексов. Так, поддержание оптимальной концентрации кислорода в плазме крови обеспечивается сердечным, дыхательным, двигательным и другими рефлексами, формирующими функциональную рефлекторную систему. Учение о функциональных системах регуляции функций было развито П. К. Анохиным (1949).
У позвоночных животных в автономной нервной системе имеется три вида синаптической передачи: электрическая, химическая и смешанная. Органом с типичными электрическими синапсами является цилиарный ганглий птиц, лежащий в глубине глазницы у основания глазного яблока. Передача возбуждения здесь осуществляется практически без задержки в обоих направлениях. К редко встречающимся можно отнести и передачу через смешанные синапсы, в которых одновременно соседствуют структуры электрических и химических синапсов. Этот вид также характерен для цилиарного ганглия птиц. Основным же способом передачи возбуждения в автономной нервной системе является химический. Он осуществляется по определенным закономерностям, среди которых выделяют два принципа. Первый (принцип Дейла) заключается в том, что нейрон со всеми отростками выделяет один медиатор. Как стало теперь известно, наряду с основным в этом нейроне могут присутствовать также другие передатчики и участвующие в их синтезе вещества. Согласно второму принципу, действие каждого медиатора на нейрон или эффектор зависит от природы рецептора постсинаптической мембраны.
В автономной нервной системе насчитывают более десяти видов нервных клеток, которые продуцируют в качестве основных разные медиаторы: ацетилхолин, норадреналин, серотонин и другие биогенные амины, аминокислоты, АТФ. В зависимости от того, какой основной медиатор выделяется окончаниями аксонов автономных нейронов, эти клетки принято называть холинергическими, адренергическими, серотоиинергическими, пуринергическими и т. д. нейронами.
Каждый из медиаторов выполняет передаточную функцию, как правило, в определенных звеньях дуги автономного рефлекса. Так, ацетилхолин выделяется в окончаниях всех преганглионарных симпатических и парасимпатических нейронов, а также большинства постганглионарных парасимпатических окончаний. Кроме того, часть постганглионарных симпатических волокон, иннервирующих потовые железы и, по-видимому, вазодилататоры скелетных мышц, также осуществляют передачу с помощью ацетилхолина. В свою очередь норадреналин является медиатором в постганглионарных симпатических окончаниях (за исключением нервов потовых желез и симпатических вазодилататоров) - сосудов сердца, печени, селезенки.
Медиатор, освобождающийся в пресинаптических терминалах под влиянием приходящих нервных импульсов, взаимодействует со специфическим белком-рецептором постсинаптической мембраны и образует с ним комплексное соединение. Белок, с которым взаимодействует ацетилхолин, носит название холинорецептора, адреналин или норадреналин - адренорецептора и т. д. Местом локализации рецепторов различных медиаторов является не только постсинаптическая мембрана. Обнаружено существование и специальных пресинаптических рецепторов, которые участвуют в механизме обратной связи регуляции медиаторного процесса в синапсе.
Помимо холино-, адрено-, пуринорецепторов, в периферической части автономной нервной системы имеются рецепторы пептидов, дофамина, простагландинов. Все виды рецепторов, вначале обнаруженные в периферической части автономной нервной системы, были найдены затем в пре- и постсинаптических мембранах ядерных структур ЦНС.
Характерной реакцией автономной нервной системы является резкое повышение ее чувствительности к медиаторам после денервации органов. Например, после ваготомии орган обладает повышенной чувствительностью к ацетилхолину, соответственно после симпатэктомии - к норадреналину. Полагают, что в основе этого явления лежит резкое возрастание числа соответствующих рецепторов постсинаптической мембраны, а также снижение содержания или активности ферментов, расщепляющих медиатор (ацетилхолин-эстераза, моноаминоксидаза и др.).
В автономной нервной системе, помимо обычных эффекторных нейронов, существуют еще специальные клетки, соответствующиепостганглионарным структурам и выполняющие их функцию. Передача возбуждения к ним осуществляется обычным химическим путем, а отвечают они эндокринным способом. Эти клетки получили название трансдукторов. Их аксоны не формируют синаптических контактов с эффекторными органами, а свободно заканчиваются вокруг сосудов, с которыми образуют так называемые гемальные органы. К трансдукторам относят следующие клетки: 1) хромаффинные клетки мозгового слоя надпочечников, которые на холинергический передатчик преганглионарного симпатического окончания отвечают выделением адреналина и норадреналина; 2) юкста-гломерулярные клетки почки, которые отвечают на адренергический передатчик постганглионарного симпатического волокна выделением в кровяное русло ренина; 3) нейроны гипоталамических супраоптического и паравентрикулярного ядер, реагирующие на синаптический приток разной природы выделением вазопрессина и окситоцина; 4) нейроны ядер гипоталамуса.
Действие основных классических меадиаторов может быть воспроизведено с помощью фармакологических препаратов. Например, никотин вызывает эффект, подобный эффекту ацетилхолина, при действии на постсинаптическую мембрану постганглионарного нейрона, в то время как сложные эфиры холина и токсин мухомора мускарин - на постсинаптическую мембрану эффекторной клетки висцерального органа. Следовательно, никотин вмешивается в межнейронную передачу в автономном ганглии, мускарин - в нейро-эффекторную передачу в исполнительном органе. На этом основании считают, что имеется соответственно два типа холинорецепторов: никотиновые (Н-холинорецепторы) и мускариновые (М-холинорецепторы). В зависимости от чувствительности к различным катехоламинам адренорецепторы делят на α-адренорецепторы и β-адренорецепторы. Их существование установлено посредством фармакологических препаратов, избирательно действующих на определенный вид адренорецепторов.
В ряде висцеральных органов, реагирующих на катехоламины, находятся оба вида адренорецепторов, но результаты их возбуждения бывают, как правило, противоположными. Например, в кровеносных сосудах скелетных мышц имеются α- и β-адренорецепторы. Возбуждение α-адренорецепторов приводит к сужению, а β-адренорецепторов - к расширению артериол. Оба вида адренорецепторов обнаружены и в стенке кишки, однако реакция органа при возбуждении каждого из видов будет однозначно характеризоваться торможением активности гладких мышечных клеток. В сердце и бронхах нет α-адренорецепторов и медиатор взаимодействует только с β-адренорецепторами, что сопровождается усилением сердечных сокращений и расширением бронхов. В связи с тем что норадреналин вызывает наибольшее возбуждение β-адренорецепторов сердечной мышцы и слабую реакцию бронхов, трахеи, сосудов, первые стали называть β1-адренорецепторами, вторые - β2-адренорецепторами.
При действии на мембрану гладкой мышечной клетки адреналин и норадреналин активируют находящуюся в клеточной мембране аденилатциклазу. При наличии ионов Mg2+ этот фермент катализирует образование в клетке цАМФ (циклического 3" ,5" -аденозинмонофосфата) из АТФ. Последний продукт в свою очередь вызывает ряд физиологических эффектов, активируя энергетический обмен, стимулируя сердечную деятельность.
Особенностью адренергического нейрона является то, что он обладает чрезвычайно длинными тонкими аксонами, которые разветвляются в органах и образуют густые сплетения. Общая длина таких аксонных терминалей может достигать 30 см. По ходу терминалей имеются многочисленные расширения - варикозы, в которых синтезируется, запасается и выделяется медиатор. С приходом импульса норадреналин одновременно выделяется из многочисленных расширений, действуя сразу на большую площадь гладкомышечной ткани. Таким образом, деполяризация мышечных клеток сопровождается одновременным сокращением всего органа.
Различные лекарственные средства, оказывающие на эффекторный орган действие, аналогичное действию постганглионарного волокна (симпатического, парасимпатического и т.п.), получили название миметиков (адрено-, холиномиметики). Наряду с этим имеются и вещества, избирательно блокирующие функцию рецепторов постсинаптической мембраны. Они названы ганглиоблокаторами. Например, аммониевые соединения избирательно выключают Н-холинорецепторы, а атропин и скополамин - М-холинорецепторы.
Классические медиаторы выполняют не только функцию передатчиков возбуждения, но обладают и общебиологическим действием. К ацетилхолину наиболее чувствительна сердечнососудистая система, он вызывает и усиленную моторику пищеварительного тракта, активируя одновременно деятельность пищеварительных желез, сокращает мускулатуру бронхов и понижает бронхиальную секрецию. Под влиянием норадреналина происходит повышение систолического и диастолического давления без изменения сердечного ритма, усиливаются сердечные сокращения, снижается секреция желудка и кишки, расслабляется гладкая мускулатура кишки и т. д. Более разнообразным диапазоном действий характеризуется адреналин. Посредством одновременной стимуляции ино-, хроно- и дромотропной функций адреналин повышает сердечный выброс. Адреналин оказывает расширяющее и антиспазматическое действие на мускулатуру бронхов, тормозит моторику пищеварительного тракта, расслабляет стенки органов, но тормозит деятельность сфинктеров, секрецию желез пищеварительного тракта.
В тканях всех видов животных обнаружен серотонин (5-окситриптамин). В мозге он содержится преимущественно в структурах, имеющих отношение к регуляции висцеральных функций, на периферии продуцируется энтерохромаффинными клетками кишки. Серотонин является одним из основных медиаторов метасимпатической части автономной нервной системы, участвующей преимущественно в нейроэффекторной передаче, и выполняет также медиаториую функцию в центральных образованиях. Известно три типа серотонинергических рецепторов - Д, М, Т. Рецепторы Д-типа локализованы в основном в гладких мышцах и блокируются диэтиламидом лизергиновой кислоты. Взаимодействие серотонина с этими рецепторами сопровождается мышечным сокращением. Рецепторы М-типа характерны для большинства автономных ганглиев; блокируются морфином. Связываясь с этими рецепторами, передатчик вызывает ганглиостимулирующий эффект. Рецепторы Т-типа, обнаруженные в сердечной и легочной рефлексогенных зонах, блокируются тиопендолом. Действуя на эти рецепторы, серотонин участвует в осуществлении коронарных и легочных хеморефлексов. Серотонин способен оказывать прямое действие на гладкую мускулатуру. В сосудистой системе оно проявляется в виде констрикторных или дилататорных реакций. При прямом действии сокращается мускулатура бронхов, при рефлекторном - изменяются дыхательный ритм и легочная вентиляция. Особенно чувствительна к серотонину пищеварительная система. На введение серотонина она реагирует начальной спастической реакцией, переходящей в ритмические сокращения с повышенным тонусом и завершающейся торможением активности.
Для многих висцеральных органов характерной является пуринергическая передача, названная так вследствие того, что при стимуляции пресинаптических терминален выделяются аденозин и инозин - пуриновые продукты распада. Медиатором же в этом случае является А Т Ф. Местом его локализации служат пресинаптические терминалы эффекторных нейронов метасимпатической части автономной нервной системы.
Выделившийся в синаптическую щель АТФ взаимодействует с пуринорецепторами постсинаптической мембраны двух типов. Пуринорецепторы первого типа более чувствительны к аденозину, второго - к АТФ. Действие медиатора направлено преимущественно на гладкую мускулатуру и проявляется в виде ее релаксации. В механизме кишечной пропульсии пуринергические нейроны являются главной антагонистической тормозной системой по отношению к возбуждающей холинергической системе. Пуринергические нейроны участвуют в осуществлении нисходящего торможения, в механизме рецептивной релаксин желудка, расслабления пищеводного и анального сфинктеров. Сокращения кишечника, возникающие вслед за пуринергически вызванным расслаблением, обеспечивают соответствующий механизм прохождения пищевого комка.
В числе медиаторов может быть гистамин. Он широко распространен в различных органах и тканях, особенно в пищеварительном тракте, легких, коже. Среди структур автономной нервной системы наибольшее количество гистамина содержится в постганглионарных симпатических волокнах. На основании ответных реакций в некоторых тканях обнаружены и специфические гистаминовые (Н-рецепторы) рецепторы: Н1- и Н2-рецепторы. Классическим действием гистамина является повышение капиллярной проницаемости и сокращение гладкой мускулатуры. В свободном состоянии гистамин снижает кровяное давление, уменьшает частоту сердечных сокращений, стимулирует симпатические ганглии.
На межнейронную передачу возбуждения в ганглиях автономной нервной системы тормозное влияние оказывает ГАМК. Как медиатор она может принимать участие в возникновении пресинаптического торможения.
Большие концентрации различных пептидов, особенно субстанции Р, в тканях пищеварительного тракта, гипоталамуса, задних корешков спинного мозга, а также эффекты стимуляции последних и другие показатели послужили основанием считать субстанцию Р медиатором чувствительных нервных клеток.
Помимо классических медиаторов и «кандидатов» в медиаторы, в регуляции деятельности исполнительных органов участвует еще большое число биологически активных веществ - местных гормонов. Они регулируют тонус, оказывают корригирующее влияние на деятельность автономной нервной системы, им принадлежит существенная роль в координации нейрогуморальной передачи, в механизмах выделения и действия медиаторов.
В комплексе активных факторов видное место занимают простагландины, которых много содержится в волокнах блуждающего нерва. Отсюда они выделяются спонтанно либо под влиянием стимуляции. Существует несколько классов простагландинов: Е, G, А, В. Их основное действие - возбуждение гладких мышц, угнетение желудочной секреции, релаксация мускулатуры бронхов. На сердечно-сосудистую систему они оказывают разнонаправленное действие: простагландины класса А и Е вызывают вазодилатацию и гипотензию, класса G - вазоконстрикцию и гипертензию.
Синапсы ВНС имеют в целом такое же строение, что и центральные. Однако отмечается значительное разнообразие хеморецепторов постсинаптических мембран. Передача нервных импульсов с преганглионарных волокон на нейроны всех вегетативных ганглиев осуществляется Н-холинергическими синапсами, т.е. синапсами на постсинаптической мембране которых расположены никотинчувствительные холинорецепторы. Постганглионарные холинергические волокна образуют на клетках исполнительных органов (желез, ГМК органов пищеварения, сосудов и т.д.) М-холинергические синапсы. Их постсинаптическая мембрана содержит мускаринчувствительные рецепторы (блокатор-атропин). И в тех и других синапсах передача возбуждения осуществляется ацетилхолином. М-холинергические синапсы оказывают возбуждающее влияние на гладкие мышцы пищеварительного канала, мочевыводящей системы (кроме сфинктеров), железы ЖКТ. Однако они уменьшают возбудимость, проводимость и сократимость сердечной мышцы и вызывают расслабление некоторых сосудов головы и таза.
Постганглионарные симпатические волокна образуют 2 типа адренергических синапсов на эффекторах – a-адренергические и b-адренергические. Постсинаптическая мембрана первых содержит a1-и a2 – адренорецепторы. При воздействии НА на a1-адренорецепторы происходит сужение артерий и артериол внутренних органов и кожи, сокращение мышц матки, сфинктеров ЖКТ, но одновременно расслабление других гладких мышц пищеварительного канала. Постсинаптические b-адренорецепторы также делятся на b1 – и b2 – типы. b1-адренорецепторы расположены в клетках сердечной мышцы. При действии на них НА повышается возбудимость, проводимость и сократимость кардиомиоцитов. Активация b2-адренорецепторов приводит к расширению сосудов легких, сердца и скелетных мышц, расслаблению гладких мышц бронхов, мочевого пузыря, торможению моторики органов пищеварения.
Кроме того, обнаружены постганглионарные волокна, которые образуют на клетках внутренних органов гистаминергические, серотонинергические, пуринергические (АТФ) синапсы.
2. учение Павлова о 1 и 2 сигнальной системах .
Сигнальная система - система условно- и безусловнорефлекторных связей высшей нервной системы животных (включая человека) и окружающего мира. Различаютпервую и вторую сигнальные системы.
Термин введен академиком И. П. Павловым.
Первая сигнальная система развита практически у всех животных, тогда как вторая система присутствует только у человека и, возможно, у некоторых китообразных. Это связано с тем, что только человек способен формировать отвлечённый от обстоятельств образ. После произнесения слова «лимон» человек может представить, какой он кислый и как обычно морщатся, когда едят его, то есть произнесение слова вызывает в памяти образ (срабатывает вторая сигнальная система); если при этом началось повышенное отделение слюны, то это работа первой сигнальной системы.
Является предметом изучения физиологии высшей нервной деятельности человека.
Вторая сигнальная система - специальный тип высшей нервной деятельности человека, система «сигналов сигналов», идущих от общей (но не одинаковой) с животными первой сигнальной системы - ощущений, представлений, относящихся к окружающему миру. Речь, как вторая сигнальная система, как семиотическая система значимостей - это «идущие в кору от речевых органов есть вторые сигналы, сигналы сигналов. Они представляют собой отвлечение от действительности и допускают обобщение, что и составляет наше личное, специально человеческое, высшее мышление, создающее сперва общечеловеческий эмпиризм, а, наконец, и науку - орудие высшей ориентировки человека в окружающем мире и в самом себе». И. П. Павлов (1932).
Мозг животного отвечает лишь на непосредственные зрительные, звуковые и другие раздражения или их следы; возникающие ощущения составляют первую сигнальную систему действительности.
В процессе эволюции животного мира на этапе становления и начального развития вида Homo sapiens произошло качественное видоизменение системы сигнализации, обеспечивающее активное и коллективное адаптивное приспособительное поведение, создавшее многообразные, принятые в группе системы сигнализации и языки: слово, по выражению И. П. Павлова, становится «сигналом сигналов». Появление второй сигнальной системы - возникновение речи и языков, сигнальных систем человека с сородичами, где условные (произвольные) сигналы индивида приобретают определенные, принятые группой значения и значимости, преобразуются в знаки языка в прямом смысле этого слова - это один из важнейших результатов многомиллионнолетней эволюции социальной жизни рода Homo, передающиеся через речевую деятельность из поколения в поколение.
В изучении В. с. с. вначале преобладало накопление фактов, характеризующих значение обобщающей функции словесных сигналов, а затем - вскрытие нервных механизмов действия слова. Установлено, что процесс обобщения словом развивается как результат выработки системы условных связей (см. Условные рефлексы); при этом имеет значение не только количество связей, но и их характер: связи, выработанные во время деятельности ребёнка, облегчают процесс обобщения. При воздействии словесных сигналов наблюдаются стойкие изменения возбудимости, большая сила, частота и длительность электрических разрядов в нервных клетках определённых пунктов коры мозга. Развитие В. с. с. - результат деятельности всей коры больших полушарий; связать этот процесс с функцией какого-то ограниченного отдела мозга невозможно. В исследованиях В.с.с. в лаборатории высшей нейродинамики и психологии высших когнитивных процессов Е. И. Бойко показана плодотворность учения И. П. Павлова о динамических временных связях В.с.с. В развитие идей И. П. Павлова и Е. А. Бойко в школе Е. А. Бойко разработана общая когнитивистская модель целостного рече-мысле-языкового процесса, найдены решения сложнейших теоретических проблем психологии в ее взаимосвязях с лингвистикой, такие как вопросы соотношения языка и речи в процессах речепроизводства и речепонимания; характер связей речи с мыслью, речи с личностью говорящего; особенности развития детской речи и др. Здесь разработаны новые методы анализа публичных выступлений (интент-анализ), позволяющий в известной мере реконструировать «картину мира» говорящего - его целевые и предметные направленности, их динамику, особенности в конфликтной ситуации, в свободных условиях общения, в публичных выступлениях и др.
Существенным резервом для дальнейших исследований остаются проблемы типологии колоссальных индивидуальных различий во взаимосвязях общего и специального типов ВНД, неокортекса и эмоционально-волевой и непроизвольной регуляции деятельности и общения, пока что слабо представленных как в физиологии ВНД, так и в психолингвистических исследованиях и в антропологической лингвистике.
Вегетативная нервная система
поддерживает гомеостаз. ВНС управляет такими висцеральными функциями, как кровообращение, пищеварение и выделение, главным образом без условного или сознательного контроля. ВНС также модулирует функцию эндокринных желез, регулирующих метаболизм. ВНС имеет сенсорные и моторные компоненты и разделяется на симпатическую и парасимпатическую системы. Первые нейроны симпатической системы расположены в промежуточных рогах тораколюмбального отдела спинного мозга; синапс со вторым набором нейронов находится в параили превертебральном симпатическом ганглии. В парасимпатической системе первые нейроны расположены либо в черепно-мозговом нерве, в автономных ядрах, либо в промежуточном роге сакрального отдела спинного мозга; синапс со вторым набором нейронов находится или в автономном ганглии (в случае черепно-мозговых нервов), или в эффекторной ткани непосредственно. ВНС имеет три главных компонента:
афферентный (центростремительный, чувствительный);
центральный объединяющий;
эфферентный.
Афферентный компонент несет информацию от нейрональных физиологических рецепторов, расположенных в концах центростремительных нервов, к спинному мозгу и более высоким областям ЦНС. Большая часть этой информации обрабатывается в пределах гипоталамуса и других нижележащих областей мозга. После обработки соответствующий сигнал посылается от ЦНС вниз по эфферентным нервам к исполнительным органам (см. рис. 8.1, 8.9), названным так потому, что они отвечают на деятельность в ЦНС.
На основе различий анатомии
и медиаторов эфферентную часть ВНС подразделяют на три системы:
парасимпатическую (холинергическую);
симпатическую (адренергическую);
неадренергическую нехолинергическую (НАНХ).
Ацетилхолин - нейромедиатор холинергической системы. - нейромедиатор, высвобождаемый из пресинаптического окончания в автономном ганглии и в окончаниях нервов в исполнительном органе. Рецепторами для ацетилхолина служат холинорецепторы, которые подразделяют на мускариновые и никотиновые.
Норадреналин - нейромедиатор адренергической системы. Другая важная составляющая ВНС - адренергическая система. До сих пор неизвестно, какой нейромедиатор использовался в этой системе первоначально - эпинефрин или норэпинефрин. Сейчас известно, что за исключением надпочечников, которые секретируют эпинефрин (адреналин), нейроме-диатором в адренергической системе является норэпинефрин.
Ацетилхолин - ганглионарный медиатор для холинергической и адренергической систем. Эфферентные нервы и для холинергической, и для адренергической систем происходят из соответствующих частей ствола мозга и спинного мозга. Эфферентные нервы образуют синапс в ганглии, расположенном вне органа, где основным нейромедиатором является АХ:
В адренергической системе ганглии находятся в цепочке вблизи спинного мозга, известной как паравертебральная симпатическая цепочка;
в холинергической системе ганглий обычно располагается внутри или вблизи эффекторного органа.
Несмотря на явное анатомическое различие
, оба типа ганглиев используют АХ как основной ганглионарный нейромедиатор, активирующий никотиновые рецепторы.
Нейромедиаторы
могут модулировать собственное высвобождение. Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут активировать пресинаптические рецепторы на нейроне, что ингибирует высвобождение самих нейромедиаторов.
Учебное видео анатомии вегетативной нервной системы (ВНС)
При проблемах с просмотром скачайте видео со страницы- Вернуться в оглавление раздела " "



